Mechanizmus IP3
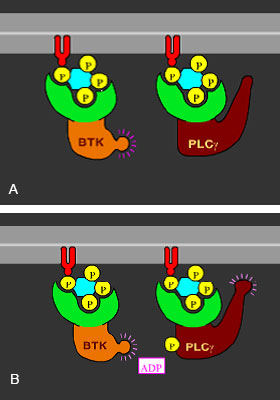
Aktivácia enzýmov IP3. A) PIP3 recrutuje tyrosinkinasu (BTK) a fosfolipasu C (PLC) B) BTK fosforyluje PLC, PLC je aktivovaná
inozitol-1,4,5-trifosfát (IP
3
) – vzniká z fosfatidylinozitolu bunkovej membrány pôsobením fosfolipázy C a zúčastňuje sa na prenose hormónového podnetu do bunky. Uvoľňuje sa z fosfatidylinozitol-4,5-bisfosfátu (PtdInsP2) v plazmatickej membráne rôznymi membránovými receptormi pre hormóny a neurotransmitery, po obsadení receptora. IP3 je aktívnou formou inozitolfosfátu ktorý zodpovedá za uvoľnenie Ca2+ a následnú stimuláciu bunky; je to druhý posol pre acetylcholín, vazopresín, substanciu P a epidermový rastovýfaktor (EGF). IP3 sa rýchlo v troch stupňoch defosforyluje na myo-inozitol. Poslednú z týchto reakcií katalyzuje myo-inozitol-1-fosfatáza (EC 3.1.3.25), ktorá je
inhibovaná lítiom. Pri syntéze myo-inozitolu de novo vzniká skôr myo-inozitol-1-fosfát ako voľný myo-inozitol, takáţo inhibícia tohto enzýmu prerušuje metabolický cyklus a zabraňuje tvorbe fosfatidylinozitolu.
Fosfatidylinozitol sa vyskytuje vo väčšom množstve ako PtdIns4P alebo PtdIns4,5P, medzi týmito formami je však rovnováha, takţe PtdIns4,5P uvoľnený odštiepením z membrány sa rýchlo dopĺňa.
Diacylglycerol (DAG) uvoľnený odštiepením z PtdIns4,5P a je podobne ako IP3, druhým poslom. Okrem iného aktivuje membránovú kinázu C, ktorá aktivuje fosforyláciou rôzne iné proteíny. DAG v PtdIns4,5P máva v polohe 2 kys. arachidonovú; keď sa kys. arachidonová hydrolýzou uvoľní z DAG, slúži tiež ako bunkový aktivátor.
inhibovaná lítiom. Pri syntéze myo-inozitolu de novo vzniká skôr myo-inozitol-1-fosfát ako voľný myo-inozitol, takáţo inhibícia tohto enzýmu prerušuje metabolický cyklus a zabraňuje tvorbe fosfatidylinozitolu.
Fosfatidylinozitol sa vyskytuje vo väčšom množstve ako PtdIns4P alebo PtdIns4,5P, medzi týmito formami je však rovnováha, takţe PtdIns4,5P uvoľnený odštiepením z membrány sa rýchlo dopĺňa.
Diacylglycerol (DAG) uvoľnený odštiepením z PtdIns4,5P a je podobne ako IP3, druhým poslom. Okrem iného aktivuje membránovú kinázu C, ktorá aktivuje fosforyláciou rôzne iné proteíny. DAG v PtdIns4,5P máva v polohe 2 kys. arachidonovú; keď sa kys. arachidonová hydrolýzou uvoľní z DAG, slúži tiež ako bunkový aktivátor.